GENOMIC
Mapping
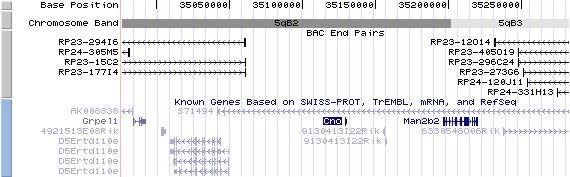
5qB2. View the map and BAC contig (data from UCSC genome browser).
Structure
(assembly 10/03)
Cno/NM_133724: 1 exons (intronless), 1,500bp, chr5:35,128,851-35,130,340.
The figure below shows the map of the known isoform (data from UCSC genome browser).

Regulatory Element
Search the 5'UTR and 1kb upstream regions (human and mouse) by CONREAL with 80% Position Weight Matrices (PWMs) threshold (view results here).
TRANSCRIPT
RefSeq/ORF
Cno/NM_133724: 1,500bp, view ORF and the alignment to genomic.
Note that alternative transcripts may exist because of differenct polyA signals.
Expression Pattern
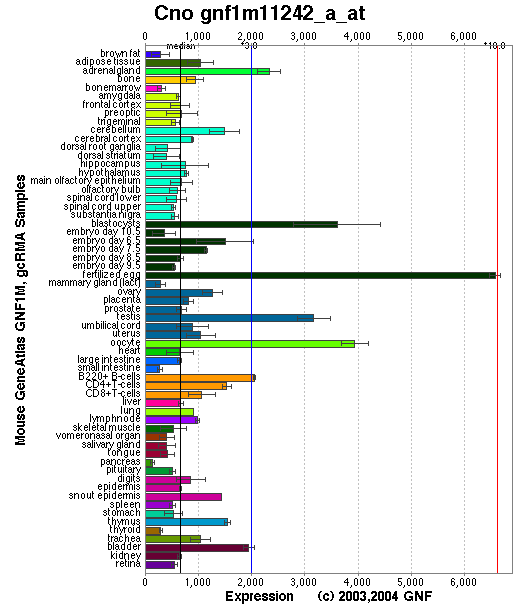
Tissue specificity: Widely expressed, highest in lung, skin, pancreas, stomach, and placenta. Two mRNA transcripts of ~1.1kb and 1.4kb are present on northern blots (Ciciotte, et al).
Affymetrix microarray expression pattern in SymAtlas from GNF is shown below.
PROTEIN
Sequence
Cappuccino protein (NP_598485): 215aa, ExPaSy NiceProt view of Swiss-Prot:Q8VED2.
Synonym: Cno.
Ortholog
| Species | Human | Rat | Fugu | Mosquito | Drosophila |
| GeneView | CNO | LOC364183 | 132443 | Q7PP24 | CG14149 |
| Protein | NP_060836 (217aa) | XP_344256 (215aa) | 140268 (210aa) | XP_315539 (184aa) | NP_648414 (169aa) |
| Identities | 77%/168aa | 93%/200aa | 37%/73aa | 28%/37aa | 24%/36aa |
View multiple sequence alignment (PDF file) by ClustalW and GeneDoc.
Domain
(1) Domains predicted by SMART:
a) low complexity: 27 - 42
b) low complexity: 80 - 93
c) coiled coil: 141-162
(2) Transmembrane domains predicted by SOSUI: none.
Motif/Site
(1) Predicted results by ScanProsite:
a) Casein kinase II phosphorylation site :
15 - 18: SteE, 87 - 90: SleE
b) N-myristoylation site :
23 - 28: GAawSG, 145 - 150: GSsvAR.
c) Cell attachment sequence :
106 - 108: RGD.
(2) Predicted results of subprograms by PSORT II:
a) N-terminal signal peptide: none
b) KDEL ER retention motif in the C-terminus: none
c) ER Membrane Retention Signals: none
d) VAC possible vacuolar targeting motif: none
e) Actinin-type actin-binding motif: type 1: none; type 2: none
f) Prenylation motif: none
g) memYQRL transport motif from cell surface to Golgi: none
h) Tyrosines in the tail: none
i) Dileucine motif in the tail: none
3D Model
(1) ModBase: none.
(2) 3D models predicted by SPARKS (fold recognition) below. View the models by PDB2MGIF.
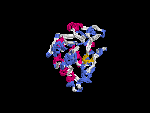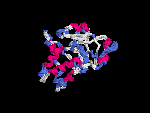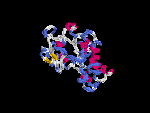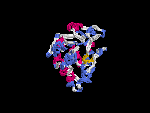
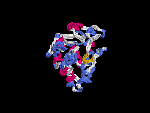
2D-PAGE
This protein does not exist in the current release of SWISS-2DPAGE.
Computed theoretical MW=23,312Da, pI=5.03 (NP_598485).
FUNCTION
Ontology
Involved in the development of lysosome-related organelles, such as melanosomes and platelet-dense granules.
Location
Cytoplasmic.
Interaction
Interacts with pallidin, muted, dysbindin, and snapin directly(Ciciotte, et al; Li, et al (2004); Starcevic, et al). The Cno protein is a subunit of the biogenesis of lysosome-related organelles complex 1 (BLOC-1), in which it interacts with the products of seven other HPS genes, mu, pa, sdy, rp, Snapap, Blos1, Blos2 (Ciciotte, et al; Falcon-Perez , et al; Li, et al (2003); Moriyama, et al; Starcevic, et al) (view diagram of BLOC-1 complex here).
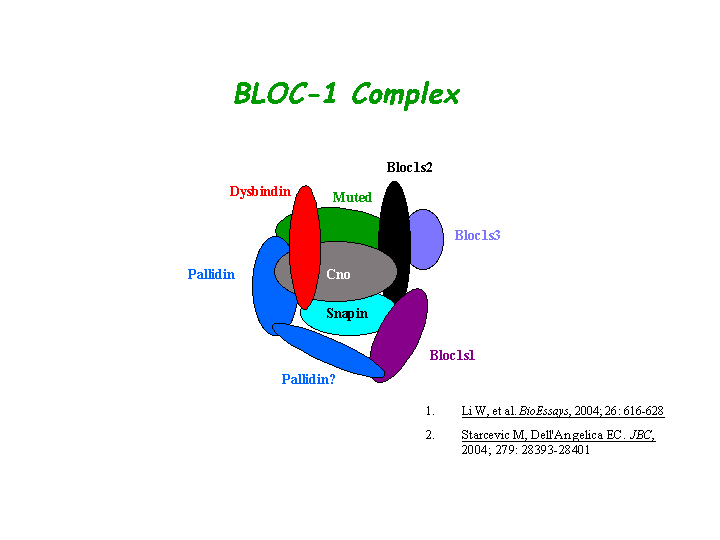{kind=link}
Cno drosophila homolog CG14149 interaction information in CuraGen interaction database.
Pathway
Nguyen, et al found that the greatest percentages of immature melanosomes were observed in the BLOC-1 mutants pa and cno, which suggests the maturation of melanosome is blocked between the multivesicular body stage and stage I (view diagram of melanosome blockage here). The cappuccino gene encodes a product involved in an AP-3-independent mechanism critical to the biogenesis of lysosome-related organelles (Gwynn, et al). (view diagram of BLOC-1 and AP-3 pathway here)
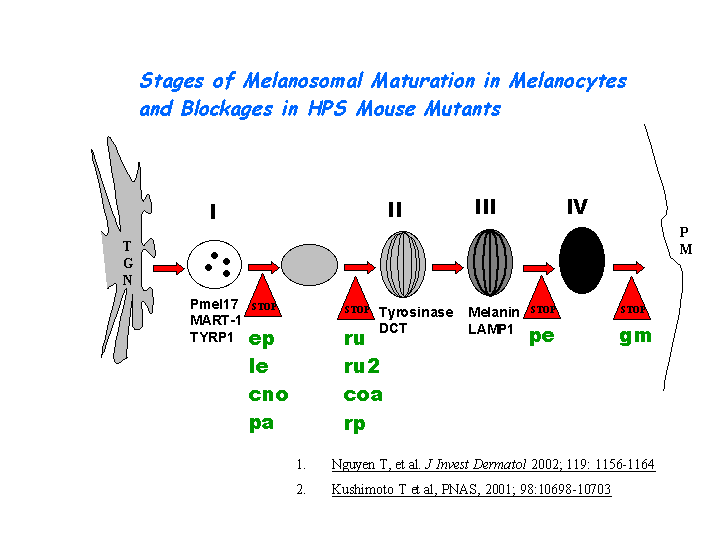{kind=link}
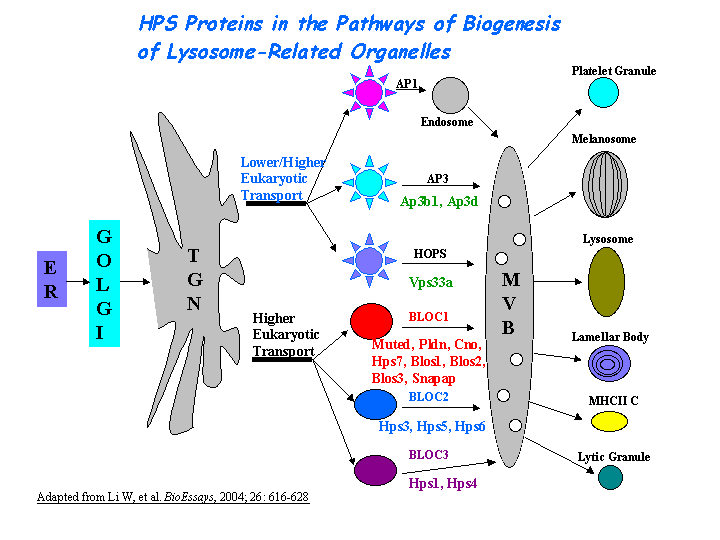{kind=link}
MUTATION
Allele or SNP
1 phenotypic allele is described in MGI:1929230.
Distribution
| Location | Genomic | cDNA | Protein | Type | Strain | Reference |
| Exon 1 | 403G~413A del | 403G~413A del | D135del 11bp | frame-shift, 207X | cno (C3H/HeJ) | Ciciotte, et al |
Effect
The deletion removes a TaqI site. Expression and subcellular localization of the protein are not affected by this mutation. The mutation does affect the stability of other subunits such as pallidin and muted of BLOC-1 complex.
PHENOTYPE
Mutation in the Cno gene is the cause of cappuccino mutant (Ciciotte, et al), a mouse model of Hermansky-Pudlak syndrome (OMIM 605695). The cno allele arose from C3H/HeJ. Homozygous mutant animals exhibit a very dilute coat and eye color due to a reduced number of melanosomes. 75% of homozygous mutant animals exhibit some form of posture or balance abnormality, with variable severity. Platelet dense bodies are markedly deficient leading to prolonged bleeding (Gwynn, et al). The phenotype is described in more detail in MGI.
REFERENCE
- Ciciotte SL, Gwynn B, Moriyama K, Huizing M, Gahl WA, Bonifacino JS, Peters LL. Cappuccino, a mouse model of Hermansky-Pudlak syndrome, encodes a novel protein that is part of the pallidin-muted complex (BLOC-1). Blood 2003; 101: 4402-7. PMID: 12576321
- Falcon-Perez JM, Starcevic M, Gautam R, Dell'Angelica EC. BLOC-1, a novel complex containing the pallidin and muted proteins involved in the biogenesis of melanosomes and platelet-dense granules. J Biol Chem 2002; 277: 28191-9. PMID: 12019270
- Gwynn B, Ciciotte SL, Hunter SJ, Washburn LL, Smith RS, Andersen SG, Swank RT, Dell'Angelica EC, Bonifacino JS, Eicher EM, Peters LL. Defects in the cappuccino (cno) gene on mouse chromosome 5 and human 4p cause Hermansky-Pudlak syndrome by an AP-3-independent mechanism. Blood 2000; 96: 4227-35. PMID: 11110696
- Li W, Rusiniak ME, Chintala S, Gautam R, Novak EK, Swank RT. Murine Hermansky-Pudlak syndrome genes: regulators of lysosome-related organelles. Bioessays 2004; 26: 616-28. PMID: 15170859
- Li W, Zhang Q, Oiso N, Novak EK, Gautam R, O'Brien EP, Tinsley CL, Blake DJ, Spritz RA, Copeland NG, Jenkins NA, Amato D, Roe BA, Starcevic M, Dell'Angelica EC, Elliott RW, Mishra V, Kingsmore SF, Paylor RE, Swank RT. Hermansky-Pudlak syndrome type 7 (HPS-7) results from mutant dysbindin, a member of the biogenesis of lysosome-related organelles complex 1 (BLOC-1). Nat Genet 2003; 35: 84-9. PMID: 12923531
- Moriyama K, Bonifacino JS. Pallidin is a component of a multi-protein complex involved in the biogenesis of lysosome-related organelles. Traffic 2002; 3: 666-77. PMID: 12191018
- Nguyen T, Novak EK, Kermani M, Fluhr J, Peters LL, Swank RT, Wei ML. Melanosome morphologies in murine models of hermansky-pudlak syndrome reflect blocks in organelle development. J Invest Dermatol 2002; 119: 1156-64. PMID: 12445206
- Starcevic M, Dell'Angelica EC. Identification of snapin and three novel proteins (BLOS1, BLOS2, and BLOS3/reduced pigmentation) as subunits of biogenesis of lysosome-related organelles complex-1 (BLOC-1). J Biol Chem 2004; 279: 28393-401. PMID: 15102850
EDIT HISTORY:
Created by Wei Li & Jonathan Bourne 06/28/2004