GENOMIC
Mapping
3q24. View the map and BAC contig (data from UCSC genome browser).
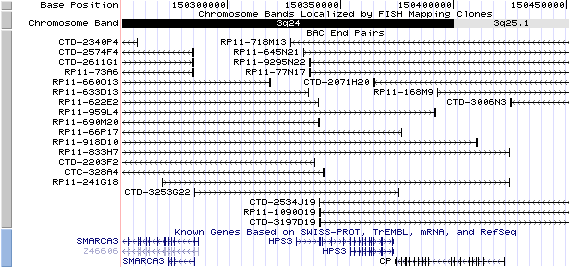
Structure
(assembly 07/03)
HPS3 (NM_032383): 17 exons, 43,936 bp, chr3:150,168,279-150,212,214.
The figure shows the structure of HPS3 gene. (data from UCSC genome browser).

Regulatory Element
Search the 5'UTR and 1kb upstream regions (seq1=human HPS3, seq2=mouse Hps3) by CONREAL with 80% Position Weight Matrices (PWMs) threshold (view results here). Conserved regions in the upstream sequences of HPS3, HPS5 and HPS6 were identified (Stanescu, et al).
TRANSCRIPT
RefSeq/ORF
HPS3/NM_032383, 4,451 bp, view ORF and the alignment to genomic.
Note that alternate splice variants exist, but their full length sequence has not been determined.
Expression Pattern
Tissue specificity: ubiquitous. Northern blot analysis demonstrated a 4.4-kb message in all tissues tested, including heart, brain, placenta, lung, liver, skeletal muscle, kidney, and pancreas. Expression was greatest in kidney, with strong bands in liver and placenta.
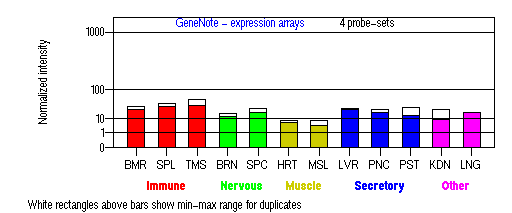
BMR: Bone marrow; SPL: Spleen; TMS: Thymus; BRN: Brain; SPC: Spinal cord; HRT: Heart; MSL: Skeletal muscle; LVR; Liver; PNC: Pancreas; PST: Prostate; KDN: Kidney; LNG: Lung. (data from GeneCards )
PROTEIN
Sequence
HPS3p (NP_115759): 1,004aa, ExPaSy NiceProt view of Swiss-Prot: Q969F9. Synonym: Hermansky-Pudlak syndrome 3 protein.
Ortholog
| Species | Mouse | Rat | Zebrafish | Drosophila |
| GeneView | coa/Hps3 | LOC310288 | ENSDARG00000015749 | CG14562 |
| Protein | NP_542365 (1,002aa) | XP_227003 (721aa) | ENSDARP00000018977 (1,001aa) | Q8MZ59 (1,190aa) |
| Identities | 79%/801aa | 64%/532aa | 47%/475aa | 17%/44aa |
View multiple sequence alignment (PDF file) by ClustalW and GeneDoc.
Domain
(1) Domains predicted by SMART:
a) ZnF NFX: 555-565
b) low complexity: 614-626
(2) Transmembrane domains predicted by SOSUI: none (predicted as a soluble protein).
Motif/Site
(1) Predicted results by ScanProsite:
a) N-glycosylation site [pattern] [Warning: pattern with a high probability of occurrence]
332-335 NLSQ, 611-614 NHSL, 854-857 NYTE, 931-934 NRTL.
b) Tyrosine sulfation site [rule] [Warning: rule with a high probability of occurrence]:
68 - 82 ayseagdYlvaieek, 402 - 416 aareedpYmdttlka, 608 - 622 fyinhslYenldeel, 848 - 862 dsladknYtedlskl.
c) cAMP- and cGMP-dependent protein kinase phosphorylation site [pattern] [Warning: pattern with a high probability of occurrence]:
284 - 287 RKyS, 459 - 462 RRqS, 469 - 472 RKdT.
d) Tyrosine kinase phosphorylation site [pattern] [Warning: pattern with a high probability of occurrence]:
119 - 126 KafrDqm.Y, 296 - 304 RfapDissY.
(2) Predicted results of subprograms by PSORT II:
a) N-terminal signal peptide: none
b) KDEL ER retention motif in the C-terminus: none
c) ER Membrane Retention Signals: KKXX-like motif in the C-terminus: KKPL
d) VAC possible vacuolar targeting motif: none
e) Actinin-type actin-binding motif: type 1: none; type 2: none
f) Prenylation motif: none
g) memYQRL transport motif from cell surface to Golgi: none
h) Tyrosines in the tail: too long tail
i) Dileucine motif in the tail: found LL at 994.
3D Model (updated 6/14)
(1) ModBase: none.
(2) 3D models predicted by SPARKS (fold recognition) below. View the models by PDB2MGIF.


2D-PAGE
This protein does not exist in the current release of SWISS-2DPAGE.
Computed theoretical MW=113,736, pI=6.01
FUNCTION
Ontology
a) Biological process: visual perception.
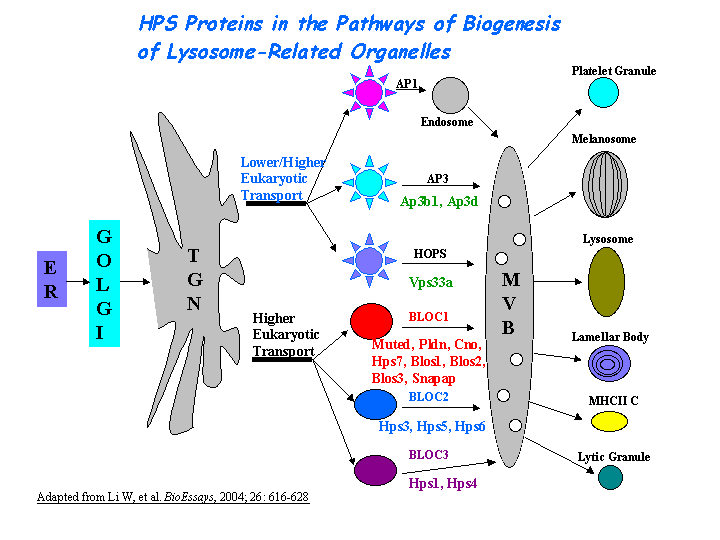
b) Plays a role in the biogenesis of lysosome-related organelles such as platelet dense granule and melanosomes (view diagram of BLOC-2 pathway here).
{kind=link}
Location
Cytoplasmic. This gene encodes a protein containing a potential clathrin-binding motif (172-176), consensus dileucine signals, and tyrosine-based sorting signals for targeting to vesicles of lysosomal lineage.
Interaction
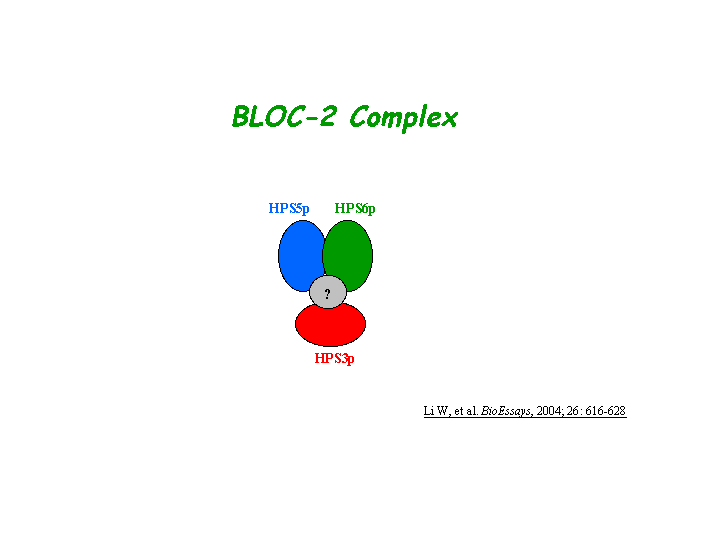
Gautum, et al reported that Hps3, Hps5, and Hps6 proteins regulate vesicle trafficking to lysosome-related organelles at the physiological level as components of the BLOC-2 (biogenesis of lysosome-related organelles complex-2) protein complex. The native molecular mass of BLOC-2 was estimated to be 340 +/- 64 kDa (view diagram of BLOC-2 complex here). BLOC-2 exists in a soluble pool and associates to membranes as a peripheral membrane protein (Di Pietro, et al). HPS3 associates with clathrin, predominantly on small clathrin-containing vesicles in the perinuclear region. This association most likely occurs directly via a functional clathrin-binding domain in HPS3 (Helip-Wooley, et al).
In HPS-1, lysosome associated membrane protein 1 (LAMP1), and LAMP3 were localized to abnormal large granules; in HPS-2, all LAMPs exhibited a normal granular expression; and in HPS-3, LAMP1, and LAMP3 exhibited a distinct less granular and more floccular pattern (Richmond, et al). Tyrosinase, tyrosinase-related protein-1 (Tyrp1), dopachrome tautomerase (Dct), LAMP1 and LAMP3 localization in HPS-3 melanocytes, as evaluated by immunocytochemistry and confocal microscopy, demonstrated a fine, floccular distribution in contrast to the coarse, granular distribution characteristic of control melanocytes (Boissy, et al).
{kind=link}
No interactions found in the CuraGen database by searching its drosophila homolog CG14562.
Pathway
Involved in early stages of melanosome biogenesis and maturation (Suzuki, et al ).
MUTATION
Allele or SNP
7 mutations deposited in HGMD.
SNPs deposited in dbSNP.
7 allelic variants described in OMIM.
Distribution
| Location | Genomic | cDNA | Protein | Type | Ethnicity | Reference |
| Exon 1 | 3904bp del | Exon 1 del | N/A | gross deletion | Central Puerto Rican |
Anikster, et al |
| Exon 3 | 727insA | 727insA | I243insA | frame-shift 247X |
? | Boissy, et al |
| Intron 5 | splicing donor +1G>C |
Exon 6 del | G324_T349 del | splicing 350X |
Ashkenazi Jewish |
Huizing, et al |
| Exon 6 | 1189C>T | 1189C>T | R397W | missense | German Swiss |
Huizing, et al |
| Intron 9 | splicing donor +2T>G |
1510_1511delGT | V504delGT | splicing 504X |
Ashkenazi Jewish |
Huizing, et al |
| Intron 13 | splicing acceptor -2A>G |
Exon 14 del | I324_Q863 del | splicing | Irish German |
Huizing, et al |
| Intron 14 | splicing donor +1G>C |
Exon 14 del | I324_Q863 del | splicing | German Swiss |
Huizing, et al |
| Intron 15 | splicing acceptor -90G>A |
2798Gins 89bp | E963ins 89bp | splicing 986X |
Irish English |
Huizing, et al |
(Numbering of genomic and cDNA sequence is based on the start codon of RefSeq NM_032383.)
Effect
Anikster, et al reported that there is a central Puerto Rican founder mutation in HPS3 (3.9 kb deletion) which causes the removal of all exon 1 and 5' partial sequence of intron 1. In non-Puerto Rican patients, splicing mutation is common. The most common splicing mutation is the splicing donor mutation on intron 5 (Huizing, et al ). Most of the mutations in HPS3 are splicing mutations which may disrupt the protein function. A compound heterozygous for the 3.9 kb deletion and a I243insA mutation results in undetectable levels of the 4.4-kb HPS3 mRNA in affected cells (Boissy, et al).
PHENOTYPE
Defects in HPS3 are the cause of Hermansky-Pudlak syndrome type 3 (HPS-3, OMIM:606118). HPS-3 is an autosomal recessive disorder, characterized by oculocutaneous albinism or ocular albinism, bleeding diathesis, and lower visual acuities. Pulmonary disease is at most mild, and no patient has a history of granulomatous colitis (Huizing, et al). Patients with HPS-3 have less severe ophthalmic manifestations than patients with HPS-1 (Tsilou, et al).
REFERENCE
- Anikster Y, Huizing M, White J, Shevchenko YO, Fitzpatrick DL, Touchman JW, Compton JG, Bale SJ, Swank RT, Gahl WA, Toro JR. Mutation of a new gene causes a unique form of Hermansky-Pudlak syndrome in a genetic isolate of central Puerto Rico. Nat Genet 2001; 28: 376-80. PMID: 11455388
- Boissy RE, Richmond B, Huizing M, Helip-Wooley A, Zhao Y, Koshoffer A, Gahl WA. Melanocyte-specific proteins are aberrantly trafficked in melanocytes of Hermansky-Pudlak syndrome-type 3. Am J Pathol 2005; 166: 231-40. PMID: 15632015
- Di Pietro SM, Falcon-Perez JM, Dell'Angelica EC. Characterization of BLOC-2, a complex containing the Hermansky-Pudlak syndrome proteins HPS3, HPS5 and HPS6. Traffic 2004; 5: 276-83. PMID: 15030569
- Gautam R, Chintala S, Li W, Zhang Q, Tan J, Novak EK, Di Pietro SM, Dell'Angelica EC, Swank RT. The Hermansky-Pudlak syndrome 3 (cocoa) protein is a component of the biogenesis of lysosome-related organelles complex-2 (BLOC-2). J Biol Chem 2004; 279: 12935-42. PMID: 14718540
- Helip-Wooley A, Westbroek W, Dorward H, Mommaas M, Boissy RE, Gahl WA, Huizing M. Association of the Hermansky-Pudlak syndrome type-3 protein with clathrin. BMC Cell Biol 2005; 6: 33. PMID: 16159387
- Huizing M, Anikster Y, Fitzpatrick DL, Jeong AB, D'Souza M, Rausche M, Toro JR, Kaiser-Kupfer MI, White JG, Gahl WA. Hermansky-Pudlak syndrome type 3 in Ashkenazi Jews and other non-Puerto Rican patients with hypopigmentation and platelet storage-pool deficiency. Am J Hum Genet 2001; 69: 1022-32. PMID: 11590544
- Richmond B, Huizing M, Knapp J, Koshoffer A, Zhao Y, Gahl WA, Boissy RE. Melanocytes derived from patients with Hermansky-Pudlak Syndrome types 1, 2, and 3 have distinct defects in cargo trafficking. J Invest Dermatol 2005; 124: 420-7.PMID: 15675963
- Stanescu H, Wolfsberg TG, Moreland RT, Ayub MH, Erickson E, Westbroek W, Huizing M, Gahl WA, Helip-Wooley A. Identifying putative promoter regions of Hermansky-Pudlak syndrome genes by means of phylogenetic footprinting. Ann Hum Genet 2009; 73: 422-8. PMID: 19523149
- Suzuki T, Li W, Zhang Q, Novak EK, Sviderskaya EV, Wilson A, Bennett DC, Roe BA, Swank RT, Spritz RA. The gene mutated in cocoa mice, carrying a defect of organelle biogenesis, is a homologue of the human Hermansky-Pudlak syndrome-3 gene. Genomics 2001; 78: 30-7. PMID: 11707070
- Tsilou ET, Rubin BI, Reed GF, McCain L, Huizing M, White J, Kaiser-Kupfer MI, Gahl W. Milder ocular findings in Hermansky-Pudlak syndrome type 3 compared with Hermansky-Pudlak syndrome type 1. Ophthalmology 2004; 111: 1599-603. PMID: 15288994
EDIT HISTORY:
Created by Wei Li: 06/16/2004
Updated by Wei Li: 08/18/2004
Updated by Wei Li: 11/24/2005
Updated by Wei Li: 12/25/2006
Updated by Wei Li: 07/31/2012
Updated by Wei Li: 06/13/2013