GENOMIC
Mapping
19qc3. View the map and BAC contig (data from UCSC genome browser).
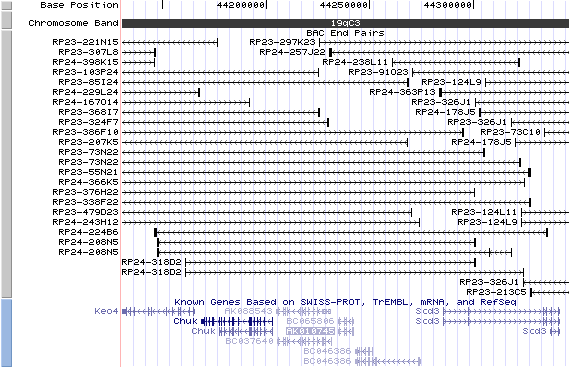
Structure
(assembly 10/03)
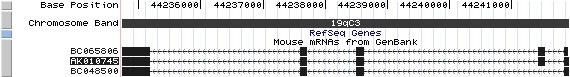
Bloc1s2/AK010745: 5 exons, 7,185 bp, chr19:44,234,734-44,241,918.
The figure below shows the structure of the Bloc1s2 gene (data from UCSC genome browser).
Regulatory Element
Search the 5'UTR and 1kb upstream regions (seq1=human BLOC1S2, seq2=mouse Bloc1s2) by CONREAL with 80% Position Weight Matrices (PWMs) threshold (view results here).
TRANSCRIPT
RefSeq/ORF
Bloc1s2/XM_193940: 887 bp, view ORF and the alignment to genomic.
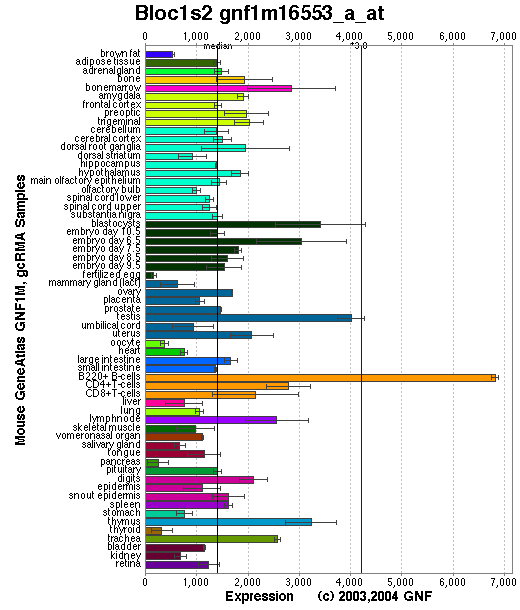
Expression Pattern
Tissue specificity: Ubiquitously expressed (by similarity).
Affymetrix microarray expression pattern in SymAtlas from GNF is shown below.
PROTEIN
Sequence
BLOC-1 subunit 2
(XP_193940): 143aa, ExPaSy NiceProt view of Swiss-Prot:Q9CWG9.
Synonym: 2410089B13 product:hypothetical protein.
Ortholog
| Species | Human | Rat | Drosophila | Mosquito | Worm |
| GeneView | BLOC1S2 | LOC293938 | CG14145 | 1276061 | 4H99 |
| Protein | NP_776170 (142aa) | XP_215245 (142aa) | NP_648427 (159aa) | XP_315366 (141aa) | NP_500967 (132aa) |
| Identities | 94%/143aa | 97%/143aa | 38%/144aa | 39%/129aa | 36%/106aa |
View multiple sequence alignment (PDF file) by ClustalW and GeneDoc.
Domain
(1) Domains of predicted by SMART:
a) low complexity: 22 - 36
b) low complexity: 128 - 142
c) coiled coil: 80-128
(2) Transmembrane domains predicted by SOSUI: none.
(3) CDD domain: KOG4559.
Motif/Site
(1) Predicted results by ScanProsite:
a) N-myristoylation site : [occurs frequently]
7 - 12: GVpaTR.
b) Protein kinase C phosphorylation site : [occurs frequently]
11 - 13: TrR,
75 - 77: SlK,
130 - 132: SkK.
c) Casein kinase II phosphorylation site : [occurs frequently]
11 - 14: TrrE,
26 - 29: TaeE,
60 - 63: TseD.
d) N-glycosylation site : [occurs frequently]
87 - 90: NISR
e) Tyrosine kinase phosphorylation site : [occurs frequently]
98 - 105: Kya.ElqpY,
131 - 137: Kkl.Eak.Y.
f) Leucine zipper pattern : [occurs frequently]
119 - 140: LeqaaykLdayskkLeakykkL
g) Tyrosine sulfation site : [occurs frequently]
57 - 71:
ltatsedYkllenmn.
(2) Predicted results of subprograms by PSORT II:
a) N-terminal signal peptide: none
b) KDEL ER retention motif in the C-terminus: none
c) ER membrane retention signals: KKXX-like motif in the C-terminus: KLEK
d) VAC possible vacuolar targeting motif: none
e) Actinin-type actin-binding motif: type 1: none; type 2: none
f) Prenylation motif: none
g) memYQRL transport motif from cell surface to Golgi: none
h) Tyrosines in the tail: none
i) Dileucine motif in the tail: none
j) DNA binding motif: Leucine zipper pattern (PS00029): found at 119.
3D Model
(1) ModBase: none.
(2) 3D models predicted by SPARKS (fold recognition) below. View the models by PDB2MGIF.
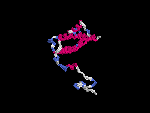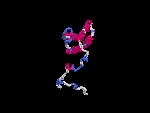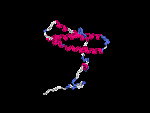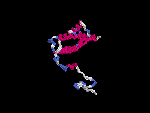
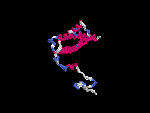
2D-PAGE
This protein does not exist in the current release of SWISS-2DPAGE.
Computed theoretical MW=16,300Da, pI=4.82.
FUNCTION
Ontology
(1) May play a role in intracellular vesicle trafficking.
(2) Protein interaction in BLOC-1.
Location
Cytoplasmic. BLOS2 also localizes to centrosomes or the pericentrosomal region, and perhaps mitochondia.
Interaction
BLOC-1 subunit 2 is a subunit of the biogenesis of lysosome-related organelles complex 1 (BLOC-1), where it resides with the products of seven other HPS genes, DTNBP1, MU, PLDN, CNO, BLOS1, BLOS3, SNAPAP (Ciciotte, et al; Falcon-Perez , et al; Li, et al; Moriyama, et al; Starcevic, et al). BLOS2 interacts with cno, muted, snapin, BLOS1, and BLOS3 within the complex ( Starcevic, et al) (view diagram of BLOC-1 complex here).
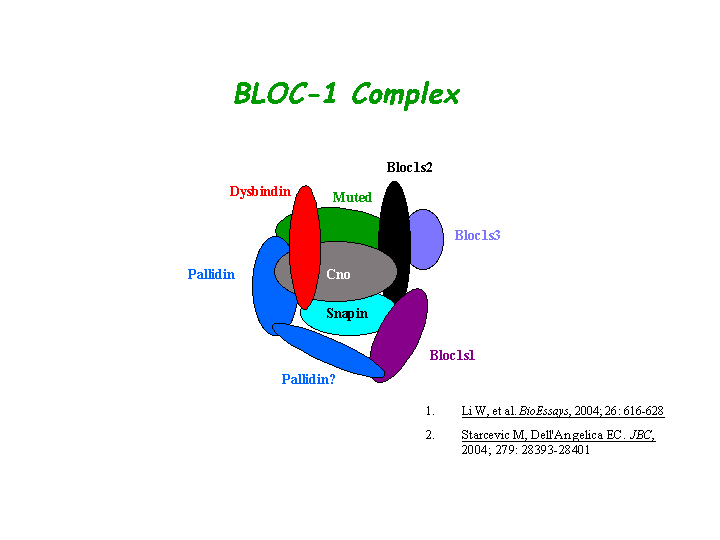{kind=link}
Two alternatively human spliced isoforms of BLOS2, which are named as Ceap-11 (isoform b) and Ceap-16 (isoform a), are co-immunoprecipitated and colocalized with gamma-tubulin (Wang, et al). Ceap-16 interacts with BRD7 (bromodomain containing protein 7), a negative regulator of cell proliferation and growth. The C-terminus of BRD7 and the central region of Ceap-16 mediated the interaction. Ceap-16 may translocate from cytoplasm to the nucleus where it selectively inhibited the transcriptional suppression activity of BRD7. Furthermore, Ceap-16, BRD7 and histone H3/H4 could form a complex and Ceap-16 did not compete with the BRD7 binding to histones, suggesting a novel function for Ceap-16 in the transcriptional regulation through BRD7 (Sun, et al). AATF/Che-1 is a coactivator of several transcription factors, including steroid hormone receptors. BLOS2 was identified as an AATF interacting protein from a rat cDNA library. BLOS2 is conserved with a high degree of homology to yeast She3p. BLOS2 functions as an adapter in processes such as protein and vesicle processing and transport, and perhaps transcription (Felten, et al). In yeast two-hybrid screen, BLOS2 was identified as a novel HIPPI (HIP-1 protein interactor) interacting protein. In co-immunoprecipitation assays, BLOS2 specifically associates with HIPPI, but not with HIP-1 (Huntingtin-interacting protein-1), suggesting that BLOS2 might play an important functional role in cancer and neurodegenerative diseases (Gdynia, et al).
BLOS2 drosophila homolog CG14145 interaction information in CuraGen interaction database.
Pathway
Involved in the development of lysosome-related organelles, such as melanosomes and platelet-dense granules (view diagram of BLOC-1 pathway here).
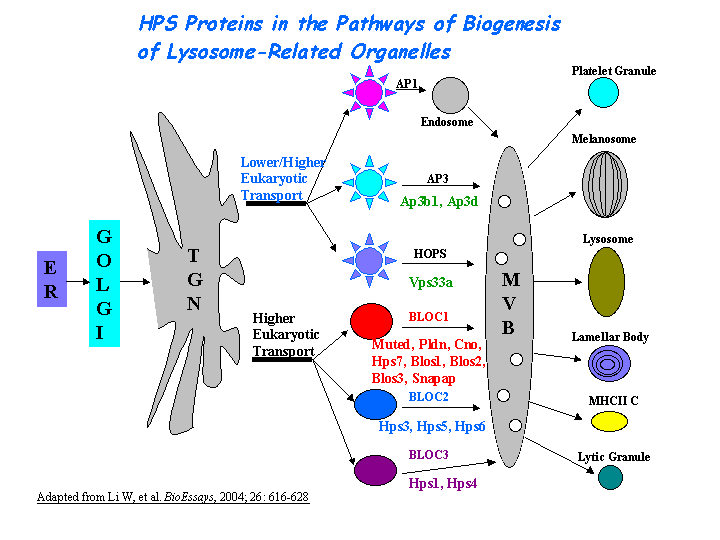{kind=link}
MUTATION
Allele or SNP
No phenotypic allele is described in MGI:1920939.
No mutation has been reported in mouse mutants.
SNPs deposited in dbSNP.
Distribution
(none)
Effect
(none)
REFERENCE
- Ciciotte SL, Gwynn B, Moriyama K, Huizing M, Gahl WA, Bonifacino JS, Peters LL. Cappuccino, a mouse model of Hermansky-Pudlak syndrome, encodes a novel protein that is part of the pallidin-muted complex (BLOC-1). Blood 2003; 101: 4402-7. PMID: 12576321
- Falcon-Perez JM, Starcevic M, Gautam R, Dell'Angelica EC. BLOC-1, a novel complex containing the pallidin and muted proteins involved in the biogenesis of melanosomes and platelet-dense granules. J Biol Chem 2002; 277: 28191-9. PMID: 12019270
- Felten A, Leister P, Burgdorf S, Uhlmann L, Scheidtmann KH. Characterization of rat BLOS2/Ceap, a putative yeast She3 homolog, as interaction partner of apoptosis antagonizing transcription factor/Che-1. Biol Chem 2007; 388: 569-82.PMID: 17552904
- Gdynia G, Lehmann-Koch J, Sieber S, Tagscherer KE, Fassl A, Zentgraf H, Matsuzawa S, Reed JC, Roth W. BLOC1S2 interacts with the HIPPI protein and sensitizes NCH89 glioblastoma cells to apoptosis. Apoptosis 2008; 13: 437-47. PMID: 18188704
- Li W, Zhang Q, Oiso N, Novak EK, Gautam R, O'Brien EP, Tinsley CL, Blake DJ, Spritz RA, Copeland NG, Jenkins NA, Amato D, Roe BA, Starcevic M, Dell'Angelica EC, Elliott RW, Mishra V, Kingsmore SF, Paylor RE, Swank RT. Hermansky-Pudlak syndrome type 7 (HPS-7) results from mutant dysbindin, a member of the biogenesis of lysosome-related organelles complex 1 (BLOC-1). Nat Genet 2003; 35: 84-9. PMID: 12923531
- Moriyama K, Bonifacino JS. Pallidin is a component of a multi-protein complex involved in the biogenesis of lysosome-related organelles. Traffic 2002; 3: 666-77. PMID: 12191018
- Starcevic M, Dell'Angelica EC. Identification of snapin and three novel proteins (BLOS1, BLOS2, and BLOS3/reduced pigmentation) as subunits of biogenesis of lysosome-related organelles complex-1 (BLOC-1). J Biol Chem 2004; 279: 28393-401. PMID: 15102850
- Sun J, Nie J, Hao B, Li L, Xing G, Wang Z, Zhou Y, Sun Q, Li G, Zhang L, He F. Ceap/BLOS2 interacts with BRD7 and selectively inhibits its transcription-suppressing effect on cellular proliferation-associated genes. Cell Signal 2008; 20: 1151-8. PMID: 18329849
- Wang Z, Wei H, Yu Y, Sun J, Yang Y, Xing G, Wu S, Zhou Y, Zhu Y, Zhang C, Zhou T, Zhao X, Sun Q, He F. Characterization of Ceap-11 and Ceap-16, two novel splicing-variant-proteins, associated with centrosome, microtubule aggregation and cell proliferation. J Mol Biol 2004; 343: 71-82. PMID: 15381421
EDIT HISTORY:
Created by Wei Li, 07/08/2004
Updated by Wei Li, 03/12/2009